Metabolisme adalah total keseluruhan reaksi yang terjadi di
dalam sel. Metabolisme terdiri atas 2 jenis, yaitu katabolisme dan anabolisme. Katabolisme adalah pemecahan molekul
kompleks menjadi lebih kecil, sederhana dan disertai pelepasan energi.
Pelepasan energi ini dapat atau tidak dapat digunakan untuk mendorong reaksi
lainnya. Anabolisme adalah sintesis
molekul kompleks dari beberapa molekul sederhana dan memerlukan sejumlah
energi. Pembahasan metabolisme tidak terlepas dari konsep termodinamika.
KONSEP
DASAR ENERGI
Hukum Termodinamika
1
Energi
tidak dapat diciptakan maupun dimusnahkan. Hal ini berarti dalam sistem hidup energi dari lingkungan harus dapat
ditangkap dan dikonversikan ke bentuk lain. Hal ini dilakukan oleh organisme fotosintetik
yang mampu menangkap energi panas matahari menjadi energi kimia (ATP dan NADP)
untuk proses biosintesis baik material sel maupun cadangan makanan (Gambar 9.1).
Organisme heterotrof juga mampu menagkap energi dari makanan menjadi energi (Gambar
9.2).
Hukum
Termodinamika 2
Pada setiap
reaksi energi dapat kehilangan kemampuannya menghasilkan kerja akibat sistem
yang tidak teratur (disorder). Pada sistem hidup
banyak sekali energi yang terbuang percuma (entropi).
Energi
bebas (DG)
Semua reaksi dapat dideskripsikan sebagai berikut
DH = DG + TDS, di mana H adalah total energi reaksi, S adalah jumlah energi yang hilang
(entropi), dan G adalah jumlah energi bebas. Reaksi di atas dapat diubah
menjadi DG = DH – TDS. Dengan demikian semakin tinggi entropi (DS), maka
semakin tidak teratur sebuah sistem (DG). Semakin rendah entropi, maka semakin
teratur sebuah sistem.
Reaksi
Oksidasi Reduksi (Redoks)
Sebagian besar reaksi di dalam sel berlangsung
dalam bentuk reaksi redoks. Reaksi redoks merupakan konsep dasar metabolisme.
Semua molekul memiliki elektron dan setiap molekul berpotensi menerima atau
mendonor elektron kepada molekul lainnya.

Potensial reaksi redoks untuk mendonasi elektron
dapat diukur. Para ilmuwan dengan susah payah akhirnya dapat menentukan nilai
potensial redoks pada kondisi standar (Eo) yang sebelumnya dianggap mustahil.
Tabel 9.1 menunjukkan daftar reaksi redoks yang umum terjadi dalam sel.
Tabel 9.1 Sejumlah reaksi redoks yang terjadi di dalam sel

PEMBAWA
ENERGI
Terdapat 3 aktivitas utama pemanfaatan energi di
dalam sel, yaitu sintesis molekul kompleks,
transportasi molekul, dan mengerakan struktur sel. Bagaimana sel
memperoleh energi untuk aktivitas tersebut? Sel memperoleh energi langsung dari
reaksi metabolisme atau mengkonversi ke molekul pembawa energi.Terdapat 2
molekul utama pembawa energi di sistem hidup, yaitu adenosin trifosfat (ATP)
dan nikotamida adenin dinukleotida (NAD+) (Gambar 9.3).
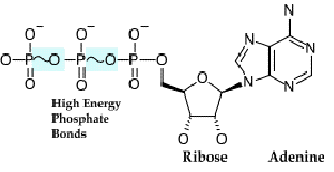

Gambar 9.3 Molekul pembawa energi ATP (atas) dan NAD+
(bawah)
ATP merupakan pembawa energi kimiawi dalam bentuk
ikatan fosfat berenergi tinggi. NAD+ merupakan pembawa hidrogen dan
elektron dan terlibat pada berbagai reaksi redoks. NAD+ dapat
mengambil dan mentransport elektron dan proton. Ketika NAD+ membawa
proton dan elektron, maka dia berubah menjadi NADH + H+. Selain ATP
dan NAD+ terdapat molekul lain sebagai pembawa energi seperti asil
fosfat. (Untuk memudahkan penulisan, maka selanjutnya NAD+ cukup
ditulis NAD, sedangkan NADH + H+ cukup ditulis NADH).
Proton
Proton atau ion hidrogen (H+) merupakan
molekul penting dalam metabolisme khususnya katabolisme. Hidrogen merupakan
elemen terkecil dan hanya memiliki proton dan elektron (tanpa neutron). Jika
kita mengambil elektron maka hidrogen berubah bentuk menjadi proton. Proten
berperan penting dalam respirasi dan reaksi enzimatik lainnya. Secara tidak
langsung proton adalah pembawa energi. Hal ini karena aktivitas translokasi
proton pada ATPase mampu menghasilkan ATP dan transportasi proton via membran
sel dapat dikopling dengan aktivitas pergerakan flagela dan silia.
ENZIM
Sebagian besar reaksi metabolisme tidak terjadi
secara spontan atau cepat, tetapi memerlukan pendorong untuk terjadinya reaksi.
Pendorong terdiri atas 2 bentuk yaitu molekul anorganik dan organik. Pendorong
anorganik disebut katalisator, sedangkan pendorong organik disebut enzim. Baik
katalisator dan enzim berperan dalam menurunkan energi aktivasi reaksi,
sehingga reaksi lebih mudah dan cepat terjadi.
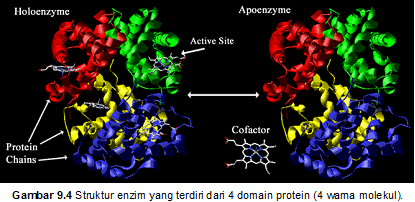
Enzim adalah protein yang berperan sebagai
katalisator metabolisme. Ukuran enzim bervariasi dari 1x104 Dalton
sampai 1x106 Dalton. Selain protein, enzim juga mengandung molekul
lain yang disebut kofaktor. Enzim lengkap dengan kofaktor disebut holoenzim, sedangkan enzim tanpa
kofaktor disebut apoenzim (Gambar 9.4).
METABOLISME
SENTRAL
Untuk dapat hidup dan berkembang, organisme harus makan. Makanan menyediakan
sumber energi dan sumber karbon untuk biosintesis komponen sel. Pada organisme
heterotrof sumber energi merupakan senyawa hidrokarbon (...-C-C-C-...). Senyawa
organik dapat monomer maupun polimer. Pada umumnya organisme heterotrof
memerlukan glukosa sebagai sumber energi dan sumber karbon. Karena glukosa
merupakan sumber energi dan sumber karbon, maka proses katabolisme glukosa
disebut metabolisme sentral. Metabolisme sentral pada organisme baik prokariota
dan eukariota adalah glikolisis dan siklus asam sitrat.
Glikolisis merupakan proses pemecahan glukosa
menjadi senyawa triosa (C3), yaitu piruvat. Siklus asam sitrat atau
siklus Krebs merupakan proses oksidasi senyawa trikarboksilat menjadi senyawa
sumber elektron atau sumber energi yang kemudian difosforilasi oksidatif
menjadi energi. Senyawa antara pada glikolisis dan siklus asam sitrat
menyediakan prekursor biosintesis asam amino.
Glikolisis
Pada eukariota glikolisis merupakan proses
pemecahan glukosa menjadi piruvat melalui jalur heksosa bisfosfat. Pada
prokariota selain jalur heksosa bisfosfat, juga terdapat jalur lain pemecahan
glukosa menjadi piruvat. Terdapat 4 jalur (pathway) glikolisis untuk katabolisme
glukosa pada prokariota, yaitu jalur Embden Meyerhoff Parnas (EMP) juga disebut
heksosa bisfosfat, jalur Heksosa Monofosfat (HMP) juga disebut fosfoketolase,
jalur Pentosa fosfat (PP), dan jalur Etner Doudoroff (ED) juga disebut
ketoglukonat. Tiga jalur yang disebut belakangan, mengubah glukosa 6-fosfat
menjadi 6-fosfo-glukonat, sedangkan jalur EMP mengubah glukosa 6-fosfat menjadi
fruktosa 6-fosfat. Keempat jalur tersebut mempunyai persamaan, yaitu memecah
heksosa (glukosa) menjadi triosa, yaitu gliseraldehid 3-fosfat (tetapi melalui
jalur berbeda), mengoksidasi triosa, menjadi asam triosa, yaitu piruvat.
Sebagian besar prokariota memproses karbohidrat
(glukosa) melalui jalur EMP dan hanya beberapa prokariota saja yang tidak
melalui jalur EMP. Sebagian prokariota mempunyai lebih dari satu jalur katabolisme
karbohidrat (glikolisis). E. coli
menggunakan jalur EMP sebagai jalur utama untuk mengonsumsi glukosa, sedangkan
jalur ED dipakai kalau sumber karbon adalah glukonat. Arkhaea Themoproteus tenax menggunakan jalur EMP
dan ED untuk mekatabolisme glikogen dan trehalosa.
Glikolisis
via Jalur EMP
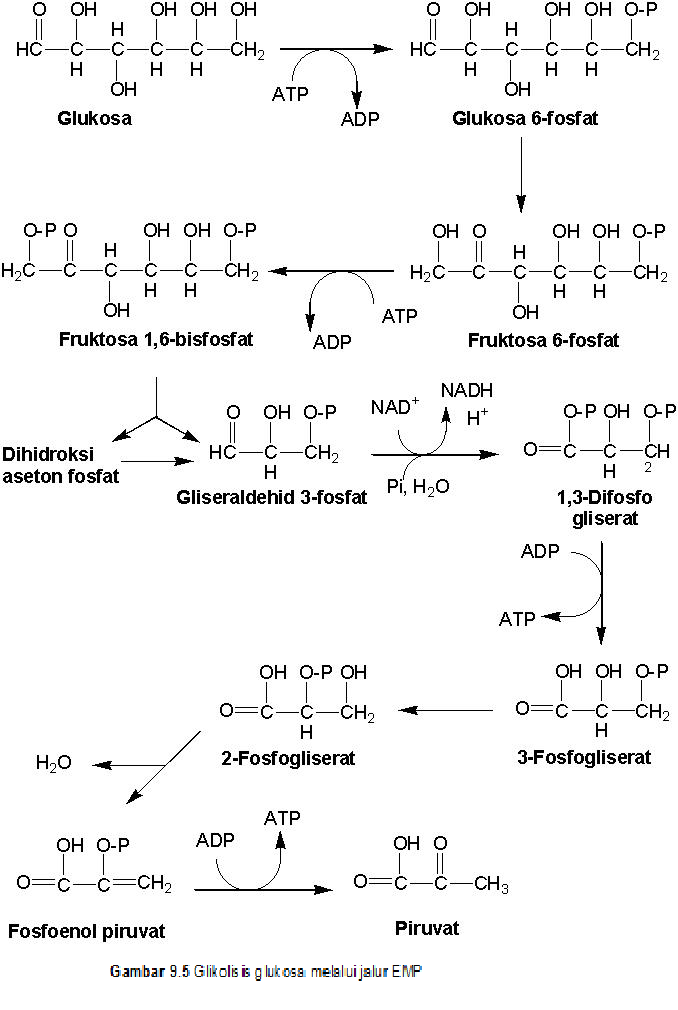
Pada jalur ini glukosa dipecah menjadi 2 piruvat
(Gambar 9.5). Jalur EMP mempunyai 4 tahapan penting katabolisme, yaitu
fosforilasi ganda heksosa, pemecahan heksosa bisfosfat menjadi 2 triosa fosfat,
dan defosforilasi triosa bisfosfat menjadi energi dan piruvat.
Tahap pertama fosforilasi
ganda heksosa dimulai dari fosforilasi glukosa menjadi glukosa 6-fosfat dengan
bantuan heksokinase. Glukosa 6-fosfat diisomerisasi (mutasi gugus karboksil
dari karboksilat menjadi keto) menjadi fruktosa 6-fosfat dengan bantuan
fosfoglukoisomerase. Selanjutnya, fruktosa 6-fosfat difosforilasi menjadi
fruktosa 1,6-bisfosfat dengan bantuan fosfofruktokinase.
Tahap kedua pemecahan
heksosa bisfosfat menjadi 2 triosa fosfat dimulai dari pemecahan fruktosa
1,6-bisfosfat menjadi gliseraldehid 3-fosfat (G3P) dan dihidroksi aseton dengan
bantuan aldolase. Dihidroksi aseton
fosfat dapat direduksi menjadi gliserol 3-fosfat dengan bantuan gliserol fosfat
dehidrogenase atau diisomerisasi menjadi G3P dengan bantuan triosa fosfat isomerase,
sehingga menghasilkan 2 triosa bisfosfat (G3P).
Tahap ketiga
defosforilasi triosa bisfosfat menjadi energi dan piruvat dimulai dari
fosforilasi G3P oleh fosfat anorganik menjadi triosa bisfosfat (1,3-difosfo
gliserat) dengan bantuan G3P dehidrogenase. Proses ini juga menghasilkan NADH
(sumber elektron bagi respirasi). 1,3-Difosfogliserat didefosforilasi
(dikopling dengan sintesis ATP) menjadi 3-fosfogliserat dengan bantuan
fosfogliserokinase. Gugus fosfat dimutasi dari posisi 3 ke posisi 2, sehingga
menghasilkan 2-fosfogliserat dengan bantuan fosfogliserat mutase. Pembentukan
ikatan rangkap (dehidrasi) antara atom C no 2 dan no 3, sehingga
2-fosfogliserat menjadi fosfoenol piruvat (PEP) dengan bantuan enolase. PEP didefosforilasi (dipasangkan
dengan sintesis ATP) menjadi piruvat dengan bantuan piruvat kinase.
Dari semua reaksi di jalur EMP terdapat beberapa
reaksi yang ireversibel (tidak balik). Reaksi yang ireversibel adalah glukosa
menjadi glukosa 6-fosfat, fruktosa 1,6 bisfosfat menjadi gliseraldehid 3-fosfat
dan dihidroksiaseton fosfat, dan fosfoenol privuvat menjadi piruvat. Hal ini
bertujuan supaya proses glikolisis tidak berlangsung bersamaan dengan proses
glukoneogenesis.
Hasil akhir adalah 2 piruvat, 2 NADH, 2 ATP.
Piruvat akan diposes lebih lanjut melalui siklus asam sitrat. NADH dioksidasi
lanjut pada respirasi atau fermentasi tergantung ada tidaknya elektron akseptor
dan enzim-enzim rantai respirasi.
Siklus Asam
Sitrat
Apa yang terjadi dengan piruvat? Piruvat akan
diurai menjadi CO2 dan menghasilkan sejumlah energi melalui
respirasi atau difermentasi. Jika difermentasi, maka piruvat direduksi menjadi
produk akhir fermentasi, seperti asam laktat dan etanol. Jika direspirasi,
piruvat dioksidasi menjadi asetil KoA dan akhirnya menjadi CO2. Pada
tulisan ini dibahas respirasi aerob (oksidasi) piruvat.
Sebelum memasuki siklus asam sitrat, piruvat dioksidasi dulu menjadi asetil KoA oleh
kompleks piruvat dehidrogenase. Kompleks piruvat dehidrogenase E. coli terdapat
di mitokondria (eukariota) dan sitoplasma (prokariota) dan terdiri atas 3
enzim, yaitu piruvat dehidrogenase (E1), dihidrolipoat transasetilase (E2), dan
dihidrolipoat dehidrogenase (E3).

Proses oksidasi piruvat menjadi asetil KoA cukup
kompleks, karena melibatkan perubahan yang terjadi pada kompleks multienzim
(Gambar 9.6). Tahapan oksidasi piruvat menjadi asetil KoA adalah sebagai
berikut. Piruvat didekarboksilasi menjadi asetaldehid aktif dan reaksi ini
dikatalisis oleh E1. Asetaldehid aktif
bereaksi dengan disulfida asam lipoat-E2 menjadi karboksil-asam lipoat. Reaksi
ini dikatalisisis E1. Transasetilasi
karboksil-asam lipoat, sehingga asam lipoat diganti KoA dan menghasilkan asetil
KoA. Reaksi ini dikatalisisis enzim 2 (E2). Sulfuridril asam lipoat-E2
dioksidasi oleh enzim 3 (E3) menjadi disulfida asam lipoat-E2. E3 tereduksi dan
proton diterima NAD , sehingga menghasilkan NADH.
Reaksi oleh kompleks piruvat dehidrogenase
merupakan reaksi tidak dapat membalik. Piruvat dehidrogenase dihambat umpan
balik oleh asetil KoA dan dipacu oleh piruvat dan AMP. AMP merupakan sinyal
bagi proses oksidasi asetil KoA menjadi NADH yang kemudian dioksidasi menghasilkan
ATP.

Gambar 9.7 Siklus TCA/Krebs/Asam Sitrat
Asetil dari asetil KoA ditransfer ke asam
oksaloasetat menjadi asam sitrat oleh sitrat sintase. Proses ini menandakan
dimulainnya siklus asam sitrat. Siklus asam sitrat mempunyai nama lain, yaitu
siklus asam sitrat dan siklus Krebs (Gambar 9.7). Reaksi asetil KoA dan
oksaloasetat menjadi sitrat yang dikatalisisis sitrat sintase. Dehidrasi sitrat
menjadi cis-akonitat yang dikatalisis akonitase. Hidrasi cis akonitat menjadi
isositrat yang juga dikatalisis akonitase. Dehidrogenasi isositrat menjadi
oksalosuksinat yang dikatalisis isositrat dehidrogenase. Dekarboksilasi
oksalosuksinat menjadi a-ketoglutarat yang dikatalisis isositrat
dehidrogenase. Dekarboksilasi dan dehidrogenasi a-ketoglutarat
menjadi suksinil KoA yang dikatalisis a-ketoglutarat dehidrogenase (perlu KoA). Konversi suksinil KoA menjadi suksinat yang
dikatalisis suksinat tiokinase. Karena ada gugus KoA yang berenergi tinggi,
maka dikopling dengan fosforilasi tingkat substrat sehingga menghasilkan ATP.
Dehidrogenasi suksinat menjadi fumarat yang dikatalisis suksinat dehidrogenase.
Penerima proton adalah FAD. Hidrasi fumarat menjadi malat yang dikatalisis
fumarase. Dehidrogenasi malat menjadi oksaloasetat yang dikatalisis malat
dehidrogenase.
Siklus asam sitrat menghasilkan 3 NADH, 1 FADH2 dan
1 ATP dari 1 molekul asetil KoA dan 2 CO2. Karena siklus asam sitrat
menghasilkan CO2, maka dimungkinkan suatu dugaan bahwa terdapat
organisme autotrof yang mampu melakukan jalur kebalikan siklus asam sitrat
untuk menambat CO2.
KATABOLISME LEMAK (b-oksidasi asam lemak)
Hampir semua lemak terpusat di membran sel. Lemak
di membran sel jarang berbentuk trigliserida, tetapi dalam bentuk lebih
kompleks, misalnya fosfolipid, glikolipid, lipoprotein, dan lainnya. Sebagian
besar lemak dalam membran sel adalah fosfolipid.
Asam lemak adalah rantai karbon metilen dengan
gugus karboksil di salah satu ujungnya. Asam lemak dapat berupa rantai metilen
lurus, atau bercabang, bahkan aromatik. Sebagian besar asam lemak dalam bakteri
sudah teresterifikasi dengan senyawa lain membentuk senyawa ester.
Sebagian prokariota (seperti Pseudomonas, Bacillus
dan E. coli) dapat tumbuh pada media yang mengandung lemak. Lemak
lebih dulu dideesterifikasi menjadi asam lemak. Asam lemak kemudian siap
mengalami oksidasi menjadi asetil KoA. Karena terjadi oksidasi di atom karbon b,
maka oksidasi tersebut dinamakan b-oksidasi.
Pada prinsipnya b-oksidasi mengurai atau memotong asam lemak menjadi
satuan (monomer) asetil KoA. Reaksi pada b-oksidasi adalah sebagai berikut (Gambar 9.8). Asam
lemak memperoleh gugus KoA, sehingga membentuk asil KoA dengan bantuan asil KoA
sintetase. Reaksi ini memerlukan energi dari ATP. Kemudian, asil KoA mengalami
dehidrogenasi, sehingga menghasilkan asil KoA tidak jenuh (mengandung ikatan
rangkap pada atom karbon b). Hidrogen yang dihasilkan, ditangkap FAD+.
Reaksi ini dikatalisis asil KoA dehidrogenase. Hidrasi ikatan rangkap pada asil
KoA tidak jenuh, sehingga menghasilkan hidroksiasil KoA. Reaksi ini dikatalisis
hidroksiasil KoA hidrolase. Dehidrogenasi gugus hidroksil dari hidroksiasil
KoA, sehingga menghasilkan gugus keto. Reaksi ini dikatalisis hidroksiasil
dehidrogenase. Hidrogen diterima oleh NAD+. Pemutusan ikatan
karbon-karbon pada posisi b akibat serangan KoA, sehingga menghasilkan asil
KoA (lebih pendek 2 atom karbon) dan asetil KoA. Reaksi ini dikatalisis b-ketothiolase.
Asil KoA kemudian mengalami perulangan reaksi-reaksi seperti di atas, sehingga
menghasilkan asil KoA yang lebih pendek 2 atom C. Asetil KoA dioksidasi menjadi
NADH, FADH, ATP, dan CO2 melalui siklus asam sitrat atau
difermentasi melalui siklus reduktif-asam sitrat menghasilkan prekursor bagi
biosintesis asam amino.
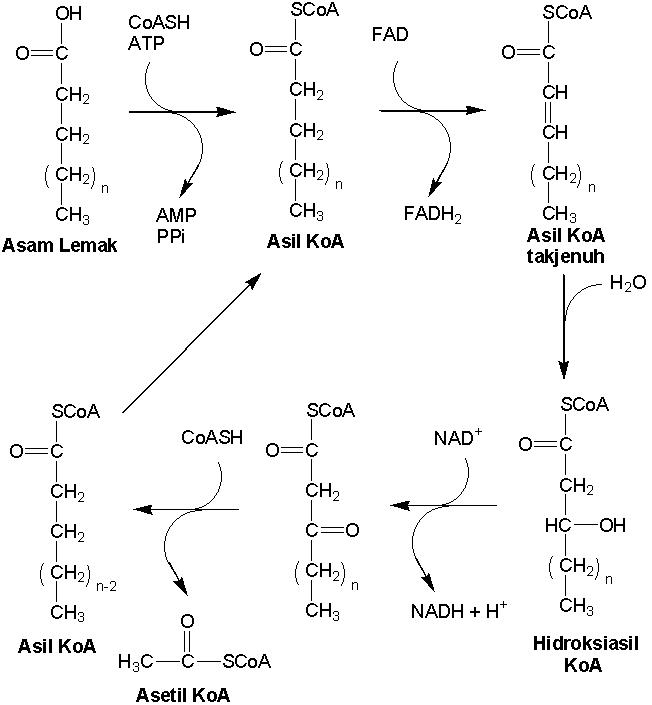
Gambar 9.8 Katabolisme lemak
Katabolisme asam amino
Asam amino dapat dipakai sebagai sumber energi. Asam amino
terlebih dulu dideaminasi menjadi asam keto. Deaminasi dapat terjadi dalam 3
model, yaitu deaminasi dengan bantuan flavoprotein oksidase, dehidrogenase, dan
transaminase. Pada proses pertama deaminasi dapat dikopling dengan pembentukan
energi. Pada proses kedua deaminasi tidak dapat langsung dikopling dengan
pembentukan energi. Akan tetapi, pada proses ketiga deaminasi dapat dikopling
dengan pembentukan energi.
Secara keseluruhan 20 asam amino didegradasi
menjadi 6 senyawa antara yang berada di metabolisme sentral, yaitu piruvat,
asetil KoA, oksaloasetat, fumarat, suksinil KoA, dan a-ketoglutarat (Gambar 9.9). Besar-kecilnya energi
yang dihasilkan tergantung pada jenis senyawa antaranya.
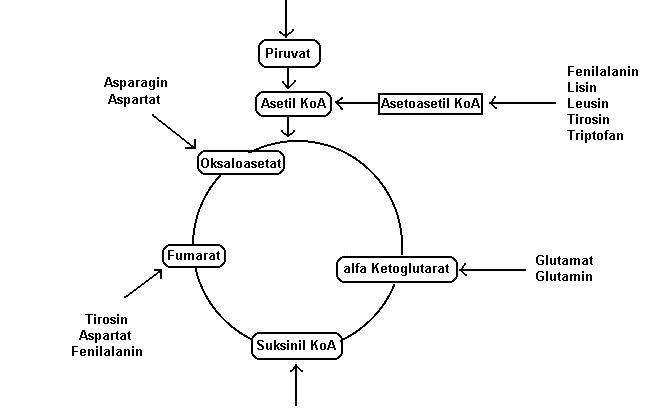

Gambar 9.9 Katabolisme protein
BIOSINTESIS
LEMAK
Biosintesis lemak dimulai dengan biosintesis
asam lemak. Prekursor biosintesis asam lemak adalah asetil KoA. Rute
biosintesis asam lemak berbeda dengan rute b-oksidasi
asam lemak (Gambar 9.10).
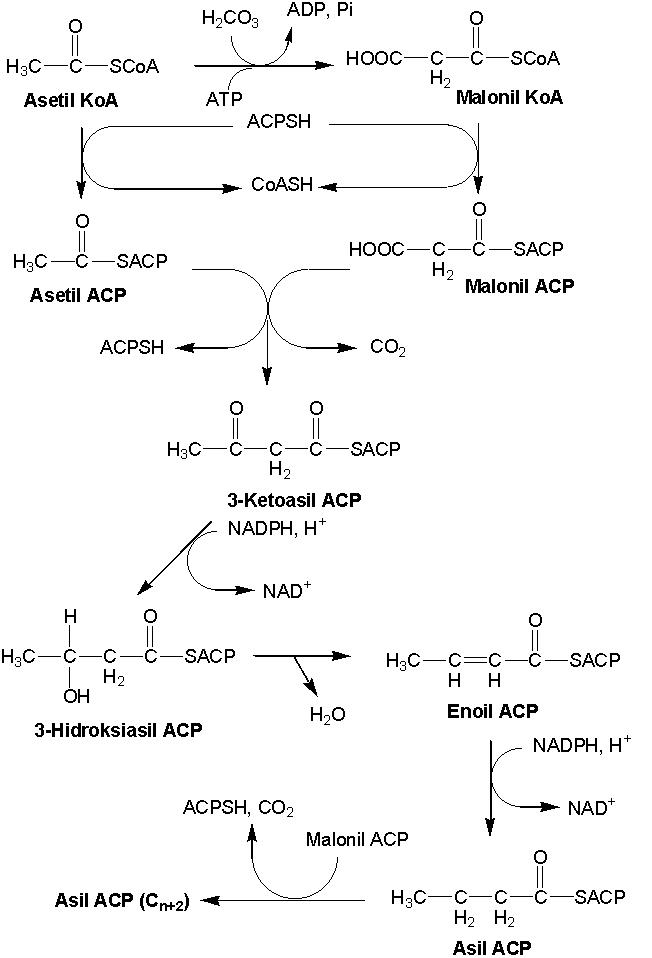
Gambar 9.10 Biosintesis asam lemak
Karboksilasi asetil KoA menjadi malonil KoA. Reaksi
ini memerlukan energi dan dikatalisis asetil KoA karboksilase. Penggantian
gugus KoA dengan gugus ACP. Reaksi ini dikatalisis malonil transasetilase. ACP
merupakan target antibiotika pantotenamida pada E. coli, sehingga mampu
menghambat biosintesis asam lemak. Di tempat terpisah, gugus KoA dari asetil
KoA diganti dengan gugus ACP, sehingga menghasilkan asil ACP. Reaksi ini
dikatalisis ketoasil ACP sintase. Asil ACP dan malonil ACP mengalami
kondensasi, sehingga menghasilkan 3-ketoasil ACP dan melepaskan CO2
dan gugus ACP. Reaksi ini dikatalisis ketoasil ACP sintase. Dehidrogensi
3-ketoasil ACP (pada C no 3) sehingga menghasilkan 3-hidroksiasil ACP. Reaksi
ini dikatalisis ketoasil ACP reduktase. Dehidrasi pada C no 3, sehingga
menghasilkan ikatan rangkap (senyawanya disebut enoil asil ACP). Reaksi ini
dikatalisis b-hidroksiasil ACP dehidrase. Dehidrogenasi pada
ikatan rangkap enoil asil ACP, sehingga menghasilkan asil (C4) ACP.
Reaksi ini dikatalisis enoil asil ACP. Kemudian, Asil ACP mengalami
pemanjangan, yaitu berkondensasi dengan malonil ACP. Reaksi ini sama dengan
reaksi asetil ACP dan malonil ACP seperti di atas. Ketika jumlah karbon dirasa
mencukupi, asil ACP ditransfer ke membran fosfolipid. Proses ini dikatalisis
gliserol fosfatase asiltransferase. Proses inkorporasi fosfat akan dijelaskan
pada tulisan berikutnya.
Biosintesis
Fosfolipid
Fosfolipid merupakan asam lemak yang terikat kovalen
dengan fosfat. Terdapat berbagai jenis fosfolipid di bakteri, tetapi yang akan
dibahas adalah fosfolipid yang umum dijumpai (Gambar 9.11). Struktur fosfolipid
dalam membran sel adalah amfibolik, yaitu satu bagian molekul bermuatan
(polar/hodrofilik) dan bagian yang lain tidak bermuatan (nonpolar/hidrofobik).
Area hidrofilik yang berisi fosfat dan hidrofobik yang berisi asam lemak
disebut kepala dan ekor.
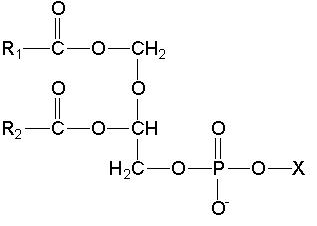
Gambar 9.11 Senyawa fosfolipid pada prokariota,
yaitu asam fosfatidil (X=H), fosfatidil serin (X=serin), fosfatidil etanolamin
(X=etanolamin), fosfatidil gliserol (X = gliserol), dan kardiolipin (X=fosfatidil
gliserol).
Beberapa sifat membran sel yang perlu diperhatikan
adalah sebagai berikut. Membran sel hanya dapat dilewati oleh air, gas, dan
molekul hidrofob kecil. Membran sel mampu melakukan kerja seluler, jika
terdapat perbedaan muatan (proton dan ion natrium) di seberang-menyeberang
membran sel. Asam lemak yang mengisi area hidrofob harus dalam kondisi cair
(fluid). Kondisi ini untuk mempertahankan fungsi membran sel. Fluiditas asam
lemak terjaga, karena adanya asam lemak tidak jenuh (titik beku asam lemak
tidak jenuh lebih rendah dibandingkan asam lemak jenuh) dan asam lemak
bercabang.
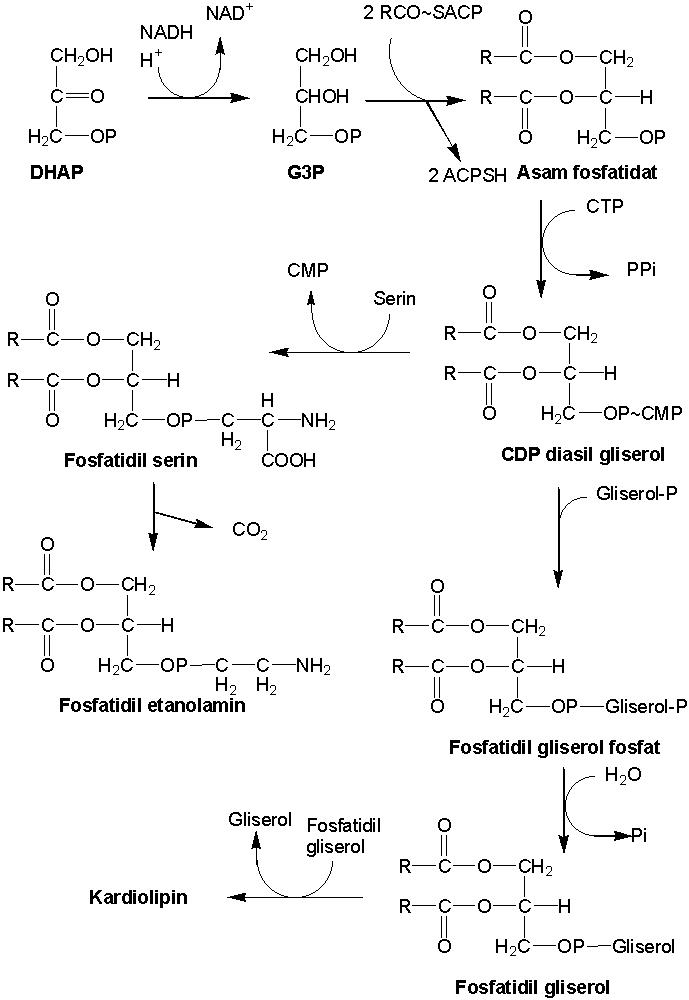
Gambar
9.12
Biosintesis fosfolipid.
Biosintesis fosfolipid dimulai dari reduksi dihidroksi
aseton fosfat (senyawa antara glikolisis) menjadi gliseraldehid 3-fosfat (G3P)
(Gambar 9.12). Dua molekul asil ACP mentransfer gugus asam lemak ke G3P
menghasilkan asam fosfatidat. Reaksi ini dikatalisis gliseraldehid 3-fosfat
asil transferase. Asam fosfatidat merupakan fosfolipid pertama yang dihasilkan.
Asam fosfatidat diproses lagi menjadi derivat fosfolipid lainnya, misalnya
fosfatidil serin, fosfatidiletanolamin, dan kardiolipin.
Asam fosfatidat kemudian bereaksi dengan sitidin
trifosfat (CTP) menghasilkan citidin difosfat diasil gliserol dan pirofosfat.
Reaksi ini dikatalisis citidin difosfat digliseride sintase. Penambahan sistein
pada citidin difosfat diasil gliserol akan menghasilkan fosfatidil serin.
Reaksi ini dikatalisis fosfatidil serin sintase. Fosfatidil serin
dekarboksilase melakukan dekarboksilasi fosfatidil serin menghasilkan
fosfatidil etanolamin. Secara terpisah, citidin difosfat diasil gliserol
bereaksi dengan gliserol fosfat menghasilkan fosfatidil gliserol fosfat. Reaksi
ini dikatalisis fosfatidil gliserol fosfat sintase. Hidrolisis fosfatidil
gliserol fosfat menghasilkan fosfatidil gliserol (dan melepaskan fosfat).
Reaksi ini dikatalisis fosfatidil gliserol fosfat fosfatase. Dua molekul
fosfatidil fosfat berekasi menghasilkan kardiolipin (difosfatidil gliserol).
Reaksi ini dikatalisis kardiolipin sintase.
BIOSINTESIS PROTEIN
Protein merupakan polipeptida dari 20 jenis asam
amino. Ke-duapuluh asam amino dapat dikelompokkan menjadi 6 kelompok
berdasarkan prekursor masing-masing (Tabel 9.2). Dapat dilihat bahwa prekursor
5 kelompok asam amino berasal dari senyawa antara pada metabolisme sentral.
Hanya histidin yang berasal dari fosforibosil pirofosfat (PRPP) atau ATP.
Meskipun sebenarnya PRPP berasal dari fosforilasi ribosa 5-fosfat dari jalur
HMP
Biosintesis Asam Amino



Tabel 9.2 Prekursor asam
amino
Prekursor |
Asam amino
|
Oksaloasetat
Piruvat
3PGA
a-Ketoglutarat
PEP & E4P
PRPP & ATP
|
Aspartat,
asaparagin, metionon, lisin, treonin, isoleusin
Alanin, valin, leusin
Serin, glisin, sistein
Glutamat, glutamin, arginin, prolin
Fenilalanin, tirosin, triptofan
Histidin
|
Biosintesis Asam Amino
Pada dasarnya sintesis asam amino merupakan reaksi
aminasi (pengikatan gugus amin) karboksilat. Gugus amin biasanya berasal dari
amonia. Akan tetapi, pengikatan gugus amin ke karboksilat dapat langsung (1
tahap) atau tidak langsung (2 tahap). Pengikatan gugus amin langsung terdapat
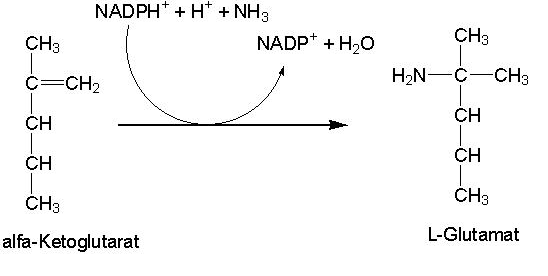
pada proses sintesis glutamat dari a-ketoglutarat
(Gambar 9.13). Reaksi ini memerlukan glutamat dehidrogenase.


Pengikatan gugus amin tidak langsung terjadi pada
proses sintesis glutamat dari a-ketoglutarat
dan memerlukan glutamat lainnya (Gambar 9.14). Pada reaksi ini gugus amin lebih
dulu diinkorporasi ke glutamat menghasilkan glutamin (dikatalisis glutamin
sintase/GS). Gugus amin ditransfer ke a-ketoglutarat
menghasilkan glutamat (dikatalisis glutamin a-oksoglutarat
transaminase atau GOGAT). Glutamin yang kehilangan amida, menjadi glutamat.
Dari kedua tipe reaksi di atas (GH dan GS-GOGAT) tampaknya
reaksi GS-GOGAT merupakan reaksi yang paling mungkin terjadi di alam. Hal ini
karena reaksi GH memerlukan kadar amonia yang tinggi di habitatnya, sedangkan
reaksi GS-GOGAT memerlukan sedikit amonia di habitatnya.
Sintesis asam amino lainnya memerlukan glutamat
sebagai sumber nitrogen (gugus amin) ke karboksilat. Reaksi ini merupakan
proses transaminasi, sehingga enzimnya disebut transaminase.
BIOSINTESIS
NUKLEOTIDA
Nukleotida adalah polimer molekul purin atau
pirimidin, ribosa atau deoksiribosa, dan fosfat (Gambar 9.15). Jika tanpa
fosfat, maka polimer tersebut dinamakan nukleosida. Jadi nukleotida adalah
nukleosida fosfat. Tiga molekul pirimidin penyusun nukleotida adalah sitosin,
timin, dan urasil dan nama nukleosidanya adalah sitidin, timidin, dan uridin.
Dua molekul purin penyusun nukleotida adalah guanin dan adenin (Gambar 9.16)
dan nama nukleosidanya adalah guanosin dan adenosin.


Biosintesis
Pirimidin
Berdasarkan asal atom karbon dan nitrogen,
pirimidin disintesis dari aspartat, amonia yang berasal dari glutamin, dan CO2
yang berasal dari karbonat (Gambar 9.17).

Sintesis nukleotida pirimidin dimulai dari
fosforilasi aminasi karbonat (Gambar 9.18). Aminasi dan fosforilasi karbonat
menjadi karbamoil fosfat. Reaksi ini dikatalisis karbamoil fosfat sintetase.
Selanjutnya, aspartat terikat pada karbamoil, sehingga menghasilkan karbamoil
aspartat (melepaskan fosfat). Reaksi ini dikatalisis aspartat
transkarbamoilase. Hidrasi karbamoil aspartat menjadi dihidroorotat. Reaksi ini

dikatalisis dihidroorotase. Dehidrogenasi
(oksidasi) dihidroorotat menjadi orotat. Reaksi ini dikatalisis dihidroorotat
dehidrogenase. Terikatnya 5-fosforibosil pirofosfat (disintesis dari
fosforilasi ribosa 5-fosfat) pada atom nitrogen dari orotat, sehingga
menghasilkan orotidin monofosfat (melepaskan pirofosfat). Reaksi ini
dikatalisis orotat fosforibosil transferase. Dekarboksilasi orotidin monofosfat
menjadi uridin monofosfat (UMP). Reaksi ini dikatalisis orotidin fosfat
dekarboksilase. Fosforilasi uridin monofosfat menjadi uridin difosfat (UDP).
Reaksi ini dikatalisis nukleosidamonofosfat kinase. Fosforilasi uridin difosfat
menjadi uridin trifosfat (UTP). Reaksi ini dikatalisis nukleosidadifosfat
kinase. Uridin trifosfat merupakan prekursor uridin.
Terikatnya amonia pada uridin trifosfat
menghasilkan sitidin trifosfat (CTP). Reaksi ini memerlukan energi/ATP. Sitidin
trifosfat merupakan prekursor sitidin. Timidin disintesis dari metilasi UTP.
Biosintesis Purin
Atom karbon dari purin berasal dari format atau
serin, glisin, dan karbon dioksida. Atom nitrogen dari purin berasal dari
aspartat, glisin, dan glutamin (Gambar 9.17).
Sintesis cincin purin dimulai dari terikatnya atom
nitrogen glutamin pada fosforibosil (berasal dari fosforibosil pirofosfat)
menghasilkan 5-fosforibosilamin (Gambar 9.19). Reaksi ini dikatalisis
fosforibosil pirofosfat amidotransferase. Tersisipnya glisin ke
5-fosforibosilamin, sehingga menghasilkan 5-fosforibosil glisinamida. Reaksi
ini memerlukan energi (ATP) dan dikatalisis fosforibosil glisinamida sintetase.
Terikatnya formil pada fosforibosil glisinamida, sehingga menghasilkan
5-fosforibosil N formilglisinamida. Reaksi ini dikatalisis fosforibosil
formilglisinamida sintetase. Pengikatan gugus NH2 dari
glutamin,sehingga menghasilkan 5-fosforibosil N formilglisinamidin. Reaksi ini
dikatalisis fosforibosil formilglisinamidin sintetase dan memerlukan energi.
Pembentukan struktur cincin pada fosforibosil N
formilglisinamidin, sehingga menghasilkan 5-fosforibosil 5-aminoimidazol.
Reaksi ini dikatalisis fosforibosil aminoimidazol sintetase. Karboksilasi
5-fosforibosil 5-aminoimidazol, sehingga menghasilkan asam 5-fosforibosil
5-aminoimidazol 4-karboksilat. Reaksi ini dikatalisis fosforibosil
aminoamidazol karboksilase. Terikatnya aspartat pada asam 5-fosforibosil
5-aminoimidazol 4-karboksilat, sehingga menghasilkan asam 5-fosforibosil 4-N
suksinokarboksamida 5-aminoimidazol. Reaksi ini dikatalisis
fosforibosilaminoamidazol suksinokarboksamida sintetase. Terlepasnya 4 karbon
yang semula berasal dari aspartat dari reaksi sebelumnya (Jadi aspartat
akhirnya hanya menyumbang nitrogen saja), sehingga
menghasilkan 5-fosforibosil 4-karboksamida 5-amidoimidazol. 
Reaksi ini dikatalisis adenilosuksinat liase.
Terikatnya formil ke 5-fosforibosil 4-karboksamida 5-amidoimidazol, sehingga
menghasilkan 5-fosforibosil 4-karboksamida 5-formamidoimidazol. Reaksi ini
dikatalisis fosforibosil amidoimidazol karboksamida formiltransferase.
Pembentukan cincin (kedua) dari 5-fosforibosil 4-karboksamida
5-formamidoimidazol, sehingga menghasilkan inosinat monofosfat (IMP). Reaksi
ini melepaskan air dan dikatalisis inosinat siklohidrolase. IMP merupakan
prekursor dari semua nukleotida purin.
REGULASI
METABOLISME
Organisme mampu mengkatalisis lebih dari seribu
reaksi kimia dalam jalur-jalur metabolisme yang saling berkaitan. Semua reaksi
dalam metabolisme dikatalisis oleh enzim. Oleh karena itu, terdapat pengaturan
(regulasi) enzim pada proses metabolisme. Regulasi enzim terdapat dalam 2
bentuk, yaitu regulasi non-kovalen (noncovalent bonding) dan regulasi
modifikasi kovalen (covalent modification). Regulasi non-kovalen adalah
terikatnya efektor oleh (biasanya) produk pada daerah alosterik (allosteric
effector) secara nonkovalen (Gambar 9.20). Regulasi modifikasi kovalen adalah
menempelnya gugus kimia (misalnya fosfat atau nukleotida) pada enzim.
Regulasi Nonkovalen
Inhibisi Umpan Balik
(feedback inhibition)
Pada reaksi biosintesis produk akhir selalu
berperan sebagai efektor alosterik negatif pada percabangan enzim (Gambar 9.21).
Penghambatan ini disebut inhibisi umpan balik. Terdapat tiga pola inhibisi
umpan balik, yaitu sederhana, kumulatif, dan terkonsentrasi (Gambar 9.22).
Jika enzim dihambat oleh 1 produk akhir saja,
maka disebut inhibisi umpan balik sederhana. Jika enzim dapat dihambat oleh 2
produk akhir, maka disebut inhibisi umpan balik kumulatif. Jika enzim dihambat
oleh 2 produk akhir secara terus-menerus sampai terjadi penghambatan, maka
disebut inhibisi umpan balik terkonsentrasi.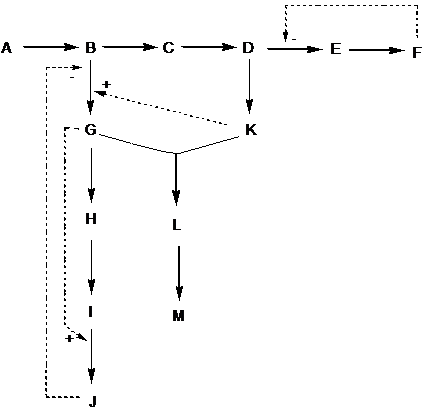
Gambar 9.21 Regulasi pada metabolisme bercabang.
Garis panah takputus menunjukkan arah reaksi, garis panah terputus menunjukkan
regulasi.

Pada inhibisi umpan balik kumulatif setiap produk
akhir tidak mempunyai efektivitas penuh menghambat aktivitas enzim. Akan
tetapi, jika kedua produk akhir bergabung, maka mampu menghambat aktivitas
enzim secara signifikan. Misalnya produk A menghambat enzim X sebesar 40%,
produk B menghambat enzim X sebesar 50%. Maka produk A dab B menghambat enzim X
sebesar 75% (bukan 40+50=90%). Akan tetapi, inhibisi umpan balik terkonsentrasi
tidak dapat dihambat oleh satu produk saja.
Regulasi Positif
Metabolisme dapat diregulasi secara positif
(dipacu). Regulasi seperti ini dapat dilakukan oleh senyawa produk antara.
Regulasi positif terdapat 2 pola, yaitu aktivasi prekursor dan regulasi positif
biasa.
Pada aktivasi prekursor enzim dipacu oleh senyawa
antara pada jalur metabolisme sama. Misalnya, enzim piruvat kinase (yang
mengkatalisis reaksi PEP menjadi piruvat ) diregulasi positif oleh fruktosa
bisfosfat (jalur glikolisis). Fruktosa bisfosfat merupakan prekursor PEP.
Pada regulasi positif biasa, jika enzim dipacu oleh
senyawa antara pada jalur metabolisme berbeda. Contoh PEP karboksilase yang
mengkatalisis PEP menjadi oksaloasetat (jalur biosinteis) di regulasi positif
oleh asetil KoA (jalur katabolisme).
Regulasi Enzimatik
Banyak reaksi yang dikatalisis enzim regulatif
merupakan percabangan reaksi. Enzim regulatif adalah enzim yang dapat
menentukan arah produk dari suatu substrat. Biasanya enzim regulatif terdiri
atas 2 jenis enzim yang mampu berinteraksi dengan substrat yang sama. Pada
metabolisme enzim regulatif biasanya ireversibel (tidak dapat mengkatalisis
balik).
Perubahan Bentuk Konformasi Enzim
Ketika
efektor berikatan dengan enzim di daerah alosterik, terjadi perubahan bentuk konformasi enzim,
sehingga mengubah kinetika enzim. Asumsikan sebuah enzim mempunyai 3 daerah
pengikatan, yaitu substrat, efektor positif, dan efektor negatif. Asumsikan
juga enzim mempunyai 2 bentuk konformasi, yaitu A dan B. Jika enzim mengikat efektor
positif (misalnya koenzim), maka enzim berbentuk konformasi A, sehingga enzim
mampu mengikat substrat dan melakukan reaksi. Jika enzim mengikat efektor
negatif (misalnya penghambat alosterik), maka enzim berbentuk konformasi B, di
mana daerah substrat mengalami perubahan, sehingga tidak mampu mengikat
substrat (Gambar 9.20).
Regulasi
Modifikasi Kovalen
Meskipun sebagian besar enzim diregulasi secara
non-kovalen (efektor alosterik), tetapi terdapat beberapa enzim atau protein
yang diregulasi secara modifikasi kovalen. Modifikasi kovalen pada enzim atau
protein biasanya dilakukan oleh gugus asetil, fosfat, metil, adenil, dan
uridil. Modifikasi kovalen biasanya merupakan perlekatan dapat pulih (tidak
permanen). Sejumlah enzim yang mengalami modifikasi kovalen tercantum dalam Tabel
9.3
Tabel
9.3 Enzim yang diregulasi secara modifikasi kovalen
Enzim
|
Modifikasi
|
Glutamin sintetase E. coli
Isositrat liase E. coli
Isositrat dehidrogenase E. coli
Histidin protein kinase sebagian besar bakteri
Protein regulator fosforilasi sebagian besar
bakteri
Sitrat liase pada Rhodopseudomonas
Protein kemotaksis E. coli
|
Adenilisasi
Fosforilasi
Fosforilasi
Fosforilasi
Asetilasi
Metilasi
|





{kind=link}
0 komentar:
Posting Komentar